У организмов, размножающихся половым путем, гены родителей перемешиваются в геноме потомства. Это позволяет естественному отбору работать не с целыми геномами, а с отдельными генами, поддерживая удачные варианты и отбраковывая неудачные. Считалось, что у бактерий отбор работает в основном на уровне целых геномов, что намного менее эффективно. Теоретически, горизонтальный обмен генами может отчасти заменить бактериям половое размножение. Однако было неясно, в какой степени это характерно для природных популяций микробов. Изучив две близкородственные популяции морских бактерий, недавно начавших приспосабливаться к разным нишам, американские биологи пришли к выводу, что адаптивная эволюция бактерий идет за счет распространения отдельных генов, а не целых геномов, то есть так же, как у организмов, размножающихся половым путем. Из этого следует, что в ходе адаптации к новым условиям горизонтальный генетический обмен успешно заменяет бактериям половое размножение.
Суть полового процесса, характерного для большинства высших организмов (эукариот), состоит в перемешивании (рекомбинации) генов (точнее, участков ДНК) разных особей. Гены, входящие в состав генома, благодаря регулярной рекомбинации перестают быть намертво связанными друг с другом. Перемешивание генетического материала позволяет естественному отбору работать на уровне отдельных генов, а не целых геномов. Это радикально увеличивает эффективность отбора и способствует быстрой адаптивной (приспособительной) эволюции. Например, если у какой-то особи в одном гене возникла полезная мутация, а в другом — вредная, то благодаря половому процессу первая из этих мутаций получает шанс распространиться в генофонде независимо от второй, а вторая, скорее всего, будет благополучно отбракована отбором независимо от первой (подробнее см. в заметке Опыты на червях доказали, что самцы — вещь полезная, «Элементы», 23.10.2009).
У бесполых организмов, практикующих клональное размножение (например, путем почкования или деления надвое), отбору приходится работать с целыми геномами. При этом судьбы всех полезных и вредных мутаций, возникающих в геноме, оказываются неразрывно связаны друг с другом. Это накладывает серьезные ограничения на адаптивные возможности организмов.
У бактерий нет настоящего полового процесса, как у эукариот, то есть регулярного перемешивания всех участков генома. Однако у них распространен горизонтальный генетический обмен (ГГО; см. Horizontal gene transfer), в ходе которого одни микробы заимствуют фрагменты ДНК у других. Заимствованные гены могут быть либо добавлены к уже имеющимся у бактерии-реципиента, либо (благодаря механизму гомологичной рекомбинации, Homologous recombination) «свой» фрагмент ДНК заменяется похожим «чужим» фрагментом.
Известно, что ГГО играет в эволюции прокариот огромную роль. Однако имеющиеся данные указывают в основном на события, которые можно условно отнести к разряду «макроэволюционных»: это сравнительно редкие случаи удачных переносов между неродственными группами микробов (см. Горизонтальный перенос генов и эволюция). Гораздо легче идентифицировать ген как заимствованный, если он получен от неродственного организма. Такой ген будет сильно отличаться от своих аналогов у ближайших родственников данной бактерии. Случаи горизонтального переноса между близкими штаммами микробов обнаружить труднее. Однако именно такой «близкородственный» ГГО, если он достаточно широко распространен в природе, мог бы отчасти заменить микробам настоящий половой процесс и повысить эффективность их адаптивной эволюции.
До сих пор неясно, играет ли ГГО между близкородственными микробами существенную роль в микроэволюционных изменениях, то есть в рутинной, повседневной адаптации бактерий к меняющимся условиям среды. Преобладает точка зрения, что отбор у бактерий всё-таки в основном клональный, действующий на уровне целых геномов. Именно такая ситуация воспроизводится, например, в долгосрочном эволюционном эксперименте Ричарда Ленски, где в пробирках эволюционируют бактерии, лишенные генов, необходимых для ГГО (см.: В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность», «Элементы», 25.03.2011).
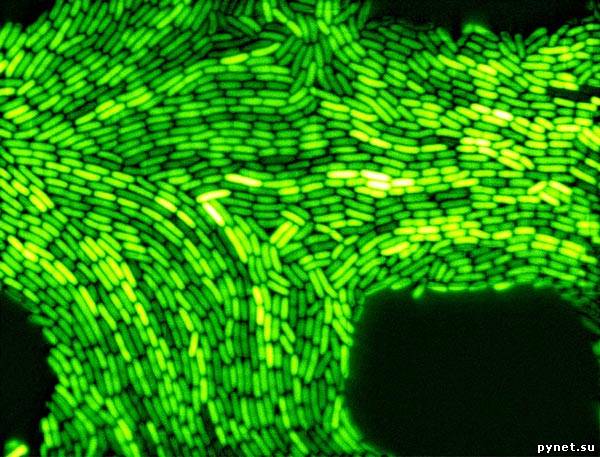
Биологи из Массачусетского технологического института (MIT) попытались прояснить вопрос о микроэволюционной роли ГГО на примере двух популяций морских планктонных бактерий Vibrio cyclitrophicus. Ранее авторы показали, что эти две популяции, обозначаемые буквами L и S, разделились недавно и в настоящее время находятся в процессе адаптации к разным экологическим нишам. Это видно по их приуроченности, соответственно, к крупным (L) и мелким (S) частицам, отфильтрованным из морской воды, и по результатам генетического анализа (см.: Hunt et al., 2008. Resource Partitioning and Sympatric Differentiation Among Closely Related Bacterioplankton, PDF, 529 Кб). По-видимому, эти две популяции приспосабливаются к жизни на разных представителях зоо- или фитопланктона.
Авторы решили выяснить, какой из двух процессов преобладает на начальных этапах экологической дифференциации — отбор отдельных генов с удачными мутациями, которые распространяются за счет ГГО, или клональный отбор, поддерживающий или отбраковывающий целые геномы. Различить эти две ситуации можно, сравнив внутри- и межпопуляционный генетический полиморфизм (вариабельность) изучаемых популяций.
Если преобладает отбор на уровне генов, две популяции должны четко отличаться друг от друга по небольшому числу генов — тех, от которых зависят экологические свойства популяций. При этом различающиеся участки генома должны иметь пониженный уровень внутрипопуляционного полиморфизма хотя бы в одной из двух популяций. Ведь на каждый такой участок действовал отбор, который поддерживал какой-то один вариант этого участка, вытесняя из генофонда другие его варианты. Напротив, те участки генома, которые у двух популяций сходны, должны быть более полиморфными, причем наборы генетических вариантов (аллелей) могут быть одинаковыми в двух популяциях (потому что отбор, связанный с приспособлением к новой нише, на них не действовал, и они могли сохранить исходный полиморфизм, накопленный предковой популяцией).
Если же в процессе расхождения популяций преобладал клональный отбор, то и уровень полиморфизма, и генетические различия между популяциями должны быть распределены по геному более равномерно.
Авторы отсеквенировали полные геномы тринадцати бактерий из популяции L и семи особей из популяции S. Анализ прочтенных геномов подтвердил первую версию: в недавней эволюции двух бактериальных популяций явно преобладал отбор на уровне отдельных генов, что было бы невозможно без интенсивного ГГО между близкородственными микробами.
Обнаружено 725 диморфных нуклеотидных позиций, в которых у всех микробов из популяции L всегда стоит какой-то один нуклеотид, а у всех микробов из популяции S — другой. Авторы назвали эти 725 позиций «экоснипами» (ecoSNPs), поскольку именно они, скорее всего, отвечают за приспособление к различающимся условиям жизни. Это видно и по функциям генов, к которым они приурочены. Экоснипы не рассеяны по геному хаотически, а сгруппированы в 11 относительно компактных кластеров. В пределах каждого из этих кластеров хотя бы у одной из двух популяций наблюдается пониженный уровень генетического полиморфизма, что свидетельствует о недавнем действии положительного отбора.
Все прочие снипы (полиморфные нуклеотидные позиции), в числе 28 744 штук, равномерно рассеяны по геному и демонстрируют сходный характер полиморфизма в обеих популяциях. Например, в какой-то позиции нуклеотид Ц может стоять у шести, а нуклеотид Г — у семи бактерий из популяции L, тогда как в популяции S нуклеотид Ц обнаружен у трех бактерий, а Г — у четырех.
Обнаруженная картина распределения нуклеотидных различий полностью соответствует предположению о том, что в ходе приспособления к различающимся условиям отбор в сочетании с ГГО (и гомологичной рекомбинацией) способствовал распространению отдельных участков ДНК с удачными мутациями, а не целых геномов, причем в остальных участках генома сохранялся исходный полиморфизм, имевшийся у предковой популяции.
Что касается гомологичной рекомбинации, то есть замены «своей» версии гена на «чужую» (заимствованную), то этот процесс характерен для так называемой центральной, или базовой (core) части генома, которая очень похожа у всех 20 изученных бактерий. В другой же, «пластичной» (flexible) части генома, в которой наборы генов и их последовательности могут сильно различаться, по-видимому, преобладали вставки чужих генов вдобавок к уже имеющимся (иногда компенсируемые утратой каких-нибудь других участков ДНК).
Еще одним подтверждением интенсивного ГГО между близкородственными микробами является то обстоятельство, что филогенетические (эволюционные) деревья, построенные для 20 изученных бактерий, оказываются разными в зависимости от того, какие фрагменты генома используются для их построения. Анализируя эти деревья, а также используя другие методы сравнительной геномики, авторы обнаружили свидетельства многочисленных недавних переносов генетического материала внутри каждой из двух популяций. В частности, удалось показать быстрое распространение целой хромосомы в популяции S. У Vibrio cyclitrophicus геном состоит из двух хромосом, обозначаемых цифрами I и II. Один из вариантов хромосомы II недавно распространился под действием отбора в популяции S (он обнаружен у 5 из 7 бактерий). Хромосома II распространялась независимо от хромосомы I, а это значит, что бактерии запросто могут обмениваться целыми хромосомами, а не только их фрагментами.
Еще один важный вывод состоит в том, что генетический обмен между популяциями в последнее время происходит редко по сравнению с обменом внутри популяций. В прошлом, когда популяции еще не окончательно разошлись по экологическим нишам, межпопуляционный обмен происходил чаще.